
La dissertation sur la race est la quatrième annexe de Sagesse des nationaux-libéraux. Elle complète le commentaire III.1.
Taxinomie
L’homme est un être vivant. L’espèce homo sapiens appartient au genre homo (seule représentante aujourd’hui), à la famille des hominidés (où elle cohabite avec les chimpanzés, les gorilles et les orang-outangs), à l’ordre des primates (qui inclut tous les singes, ainsi que les lémuriens), à la classe des mammifères, au sous-embranchement des vertébrés, à l’embranchement des chordés, au règne animal, au domaine des eucaryotes, au monde vivant.
La classification des êtres vivants est fondée exclusivement sur leur patrimoine héréditaire, leur génotype, en aucune façon sur leur phénotype, bien que ce fût seulement par celui-ci que l’on pouvait connaître celui-là avant que l’on pût faire l’analyse directe de l’ADN.
La division des espèces en races est absolument générale. Elle n’est pas réservée aux animaux domestiques, chiens ou chats, ni aux animaux d’élevage comme les chevaux, les bovins, etc., bien que pour ces deux catégories la sélection artificielle réalisée par l’homme ait multiplié les races bien davantage que ne l’a fait la sélection naturelle pour les espèces sauvages. On trouve, par exemple, quatre races de chimpanzés – espèce la plus proche de la nôtre –, deux races de gorilles, trois races d’orangs-outangs, trente-huit races de loups, quarante-cinq races de renards, vingt-deux races de ratons-laveurs, neuf races de girafes, deux races de kangourous… et donc cinq races d’hommes : caucasoïdes, mongoloïdes, congoïdes, capoïdes et australoïdes.
L’usage courant de parler pour les races humaines de « blancs », de « jaunes » et de « noirs » est triplement inadéquat.
Primo, il n’y a pas trois races humaines, mais cinq. Il y a trois races de « noirs » : les congoïdes, les capoïdes et les australoïdes, lesquelles n’ont en réalité rien à voir entre elles. Certes, les capoïdes, qui tirent leur nom de la province du Cap en Afrique du sud et qui sont représentés par les Boschimans et les Hottentots, sont aujourd’hui très métissés de congoïdes, mais ils formaient originellement une race sans rapport avec la leur. Quant aux australoïdes, représentés par les aborigènes australiens, les Mélanésiens, comme les Papous et les Canaques, les Négritos du Sud-Est asiatique, les Veddas de Ceylan (aucun rapport avec les Védas, livres sacrés de l’hindouisme !), les aborigènes de l’Inde, enfin les Aïnous, aborigènes du Japon, ils forment également une race indépendante du même niveau taxinomique que les congoïdes et les trois autres races humaines.
Secundo, ces couleurs conventionnelles sont trompeuses. Caucasoïdes ou non, les hommes n’ont jamais la peau blanche, à l’exception des albinos, et on en trouve dans toutes les races ; les albinos congoïdes ont la peau parfaitement blanche, comme ceux des autres races. Nul homme non plus n’a la peau noire ; les congoïdes ont la peau brune, même ceux qui ont la peau la plus foncée et qui sont notamment au Sénégal ou au Mali, et cela contraste avec leurs cheveux et leur barbe, qui sont, eux, vraiment noirs. Les mongoloïdes ont souvent la peau jaune, il est vrai, mais c’est très loin d’être toujours le cas ; beaucoup de Japonais ont la peau aussi claire que les Français ; et les populations du Sud-Est asiatique, en dehors du Vietnam, de même que les Mérinas de Madagascar, venus d’Indonésie, ont la peau brune et non jaune – ce qui, il est vrai, est probablement dû à un métissage ancien avec les aborigènes australoïdes.
Tertio, ces termes ont le grave inconvénient de faire uniquement référence à la couleur de la peau. Celle-ci a beau apparaître en premier lieu sous le regard et avoir en conséquence un important rôle social, elle est négligeable sur le plan biologique en tant que caractère racial. La réduction de la race à la couleur de la peau est une ânerie scientifique et une arme de désinformation pour ceux qui nient absurdement l’existence des races humaines ou qui, s’ils l’admettent à contre-cœur, soutiennent bien à tort que les différences raciales ne seraient que peu de chose.
La couleur de la peau est d’autant moins pertinente pour la taxinomie raciale que la pigmentation ne dépend pas seulement des gènes, mais aussi de l’exposition aux rayons du soleil. Les nouveaux-nés de toutes les races, tout juste sortis du ventre de leur mère, ont toujours la peau claire. Un pur nordique, blond aux yeux bleus, à la peau normalement très claire, peut l’avoir presque aussi foncée que certains congoïdes s’il a bronzé pendant des dizaines d’heures sous un soleil éclatant.
Les races ou sous-espèces ne sont nullement homogènes, loin s’en faut ; elles sont au contraire subdivisées à leur tour en de nombreuses sous-races (terme de taxinomie qui n’a rien de péjoratif), lesquelles sont en général mélangées intimement au sein d’une même population. Par exemple, pour la race caucasoïde, en France, on trouve les sous-races alpine, méditerranéenne et nordique, qui semblent se rapporter respectivement aux trois couches successives qui ont formé la population française, les chasseurs ouest-européens du paléolithique étant plutôt alpins, les paysans anatoliens du néolithique, plutôt méditerranéens, les conquérants indo-européens de l’âge du bronze, plutôt nordiques – ces trois couches représentant respectivement 15%, 45% et 40% du fonds génétique des Français de sang.
La « race aryenne » dont on parlait autrefois était un autre nom donné à la sous-race nordique de la race caucasoïde. Du reste, l’emploi du même mot, « race », dans deux acceptions, y compris par les spécialistes, pour désigner deux niveaux de la taxinomie est regrettable, étant une source de confusion. Il vaut mieux réserver le mot « race » aux sous-espèces et parler de « sous-races » pour les subdivisions de la race.
Paralogismes
La race n’est pas un concept dépassé qui remonterait au XIXe siècle. D’abord, la notion de race est en réalité plus ancienne, puisqu’elle a été définie au XVIIIe siècle par Carl von Linné, savant suédois qui a élaboré la première classification des êtres vivants (1758). Ensuite, la connaissance scientifique est un processus cumulatif. Les savants du XIXe siècle ont perfectionné la raciologie, l’étude des races, notamment pour notre espèce, homo sapiens. Le fameux naturaliste anglais Charles Darwin a fait de la race l’élément central de l’évolution dans son fameux livre de 1859, « On the origin of species » (titre qu’il faut traduire correctement : « De la genèse des espèces », et non : « De l’origine des espèces », comme on le fait d’ordinaire), puisqu’il définissait la sélection naturelle, dans le titre complet de son ouvrage (« On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life »), comme la « conservation des races avantagées dans la lutte pour la vie ». Au XXe siècle, le grand anthropologue américain Carleton Coon a établi la classification définitive des races humaines (1965). L’analyse de l’ADN n’a fait ensuite que confirmer ces analyses.
La terminologie adoptée par Coon renvoie conventionnellement chacune des cinq races à une région de référence : Caucase, Mongolie, Congo, province du Cap en Afrique du sud, Australie, sans que ces régions fussent nécessairement le foyer de dispersion de la race en question. Ni qu’elles fussent, avant les grandes découvertes et la colonisation qui a suivi, le centre de gravité de la répartition géographique de la race ; ce n’est le cas que pour le Caucase et le Congo, mais ce ne l’est ni pour la Mongolie ou l’Australie, ni même pour la province du Cap.
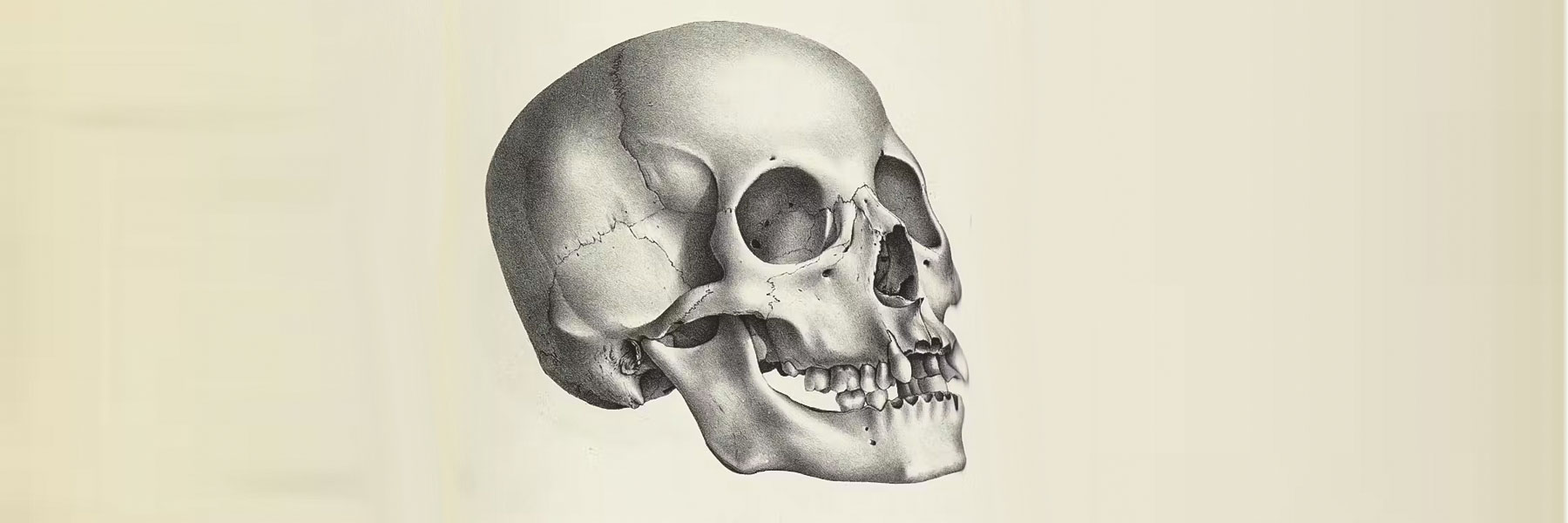
Il est absurde de reprocher aux anthropologues de mesurer les crânes humains et de calculer leur indice céphalique, rapport de la largeur à la longeur du crâne, pour distinguer dolichocéphales, mésocéphales et brachycéphales. La science repose sur la mesure, ici sur celle des squelettes, dont les os du crâne. Avant l’analyse de l’ADN et la paléogénétique, qui sont des disciplines récentes, on n’avait pas d’autre moyen d’étudier les fossiles.
Précisons que l’haplogroupe paternel, qui caractérise le seul chromosome Y, soit un sur quarante-six, ne représente qu’une très petite partie du génome (100 gènes sur 20.000) et qu’il ne suffit pas à délimiter la race puisqu’il y en a de multiples dans une même race. De surcroît, il est particulièrement susceptible de se diffuser par métissage, les conquérants ou les marchands ayant une tendance prononcée à prendre femme dans le pays où ils s’installent. Par exemple, l’haplogroupe R1b, majoritaire en France et qui nous vient des conquérants indo-européens, était à l’origine purement caucasoïde, mais on le retrouve aujourd’hui dans certaines régions d’Afrique noire.
Dans quelque domaine que ce soit, toute classification repose sur certains critères, et sur eux seulement, à l’exclusion de tous les autres, en sorte qu’un élément ou un individu peut être plus proche à d’autres égards et selon d’autres critères d’un individu d’une autre catégorie que d’un individu de la même catégorie. C’est le paradoxe de la taxinomie. Par exemple, dans une bibliothèque où les livres sont classés par genre, un essai et un roman écrits dans la même langue par le même auteur auront plus d’affinités, au moins pour ces deux critères, et probablement pour d’autres, que deux essais ou deux romans écrits dans deux langues différentes par deux auteurs différents. De même, dans l’espèce humaine, une femme jeune et mince de race caucasoïde ressemblera plus à une autre femme également jeune et mince, mais de race mongoloïde, qu’à un homme vieux et corpulent de sexe masculin, fût-il de race caucasoïde comme elle. Le classement selon la race ne recoupe évidemment pas le classement selon le sexe ou selon l’âge.
C’est encore un paralogisme d’invoquer la diversité des groupes sanguins au sein de chaque race pour nier l’existence de celles-ci. La classification raciale ne se fonde pas sur ces caractères, bien que la proportion de ces groupes varie selon les races. La meilleure preuve en est que le système ABO dépasse la barrière des espèces et qu’on le retrouve chez les singes. Il vaudrait mieux, pour un homme de groupe O, recevoir le sang d’un chimpanzé du même groupe, que d’un autre homme d’un groupe différent, fût-il de sa famille et de sa race !
Évacuons aussi le paradoxe de la continuité. De même que le spectre lumineux présente un continuum de couleurs, ce qui ne signifie pas que les couleurs n’existent pas, on trouve un continuum de populations entre les races. Ainsi, du Sin-kiang à la Bulgarie, il y a tous les stades intermédiaires entre mongoloïdes et caucasoïdes ; de même, du Liban au Kénya, en passant par la Corne de l’Afrique – Éthiopie, Érythrée, Somalie –, entre caucasoïdes et congoïdes. Ces transitions progressives sont dues à des métissages fort anciens.
Pour autant, la classification des races n’a rien d’arbitraire. Elle repose sur l’évolution génétique des populations préhistoriques réparties sur tout l’Ancien Monde et qui ne communiquaient pas ou très peu entre elles, étant observé que le cheval et le chameau de Bactriane n’ont été domestiqués qu’en -3000 et que les périodes glaciaires ont établi des barrières difficilement franchissables entre des populations éloignées. La seule question, non tranchée à ce jour, est de savoir si les races humaines se sont dissociées il y a un million d’années ou davantage, thèse dite polycentrique soutenue notamment par Carleton Coon, ou si elles l’ont fait il y a seulement 300.000 ans, thèse dite monocentrique, laquelle suppose que l’homme moderne ait exterminé partout les populations archaïques.
Évidemment, le fait que les individus de races différentes puissent avoir des enfants ne signifie pas que les races n’existent pas, mais simplement qu’elles ne sont pas au niveau taxinomique de l’espèce. En effet, c’est l’absence d’interfécondité qui permet, par définition, de distinguer des espèces voisines. Puisque les individus des diverses races appartiennent à la même espèce, ils sont donc interféconds et peuvent se croiser s’ils ne sont pas de même sexe. Un métis est un individu dont les ancêtres appartiennent à deux ou plusieurs races, et non à une seule. Il peut être aussi dit plus précisément biracial ou pluriracial, par opposition à un individu d’une seule race, uniracial.
La pureté raciale absolue n’est pas nécessaire pour être qualifié d’uniracial, dès lors que les races secondaires n’ont laissé que des traces négligeables. Pour une définition rigoureuse, on peut adopter la règle du seizième : un individu sera tenu pour uniracial, et non comme métis, si la part des autres races que la race principale dans son patrimoine héréditaire est inférieure à un seizième, soit l’équivalent de la contribution génétique d’un arrière-arrière-grand-parent ou trisaïeul.
Sur un total de huit milliards d’hommes, la population mondiale compte au moins un milliard de métis, qui sont surtout en Amérique du sud, en Amérique centrale, aux États-Unis d’Amérique, en Afrique du nord, dans la Corne de l’Afrique, en Afrique du sud, à Madagascar et dans les autres îles de l’Océan Indien, en Turquie et dans les autres pays turciques – où la population est presque entièrement métisse, hormis les minorités ethniques –, en Russie, dans le sous-continent indien et dans le sud-est de l’Asie.
Caractéristiques
L’égalité n’étant pas dans la nature, les races ne sauraient être égales. C’est ainsi que le QI moyen des congoïdes d’Afrique noire est de 70, inférieur de 15 points à celui des caucasoïdes d’Afrique du nord, du reste métissés de congoïdes, et de 30 points à celui des caucasoïdes d’Europe. Les congoïdes des États-Unis d’Amérique, qui sont en réalité métissés de caucasoïdes à 25%, ont un QI moyen inférieur de 15 points à celui des caucasoïdes du même pays. Les australoïdes ont un QI moyen inférieur de 40 points à celui des caucasoïdes occidentaux. Il est certain que ces différences de phénotype sont imputables essentiellement aux différences de génotype.
Inégalité des races humaines, donc, ainsi que le proclamait Arthur de Gobineau dans son fameux Essai de 1855, ouvrage monumental et magnifique, quoique non scientifique, ou plutôt pré-scientifique, l’état des connaissances étant alors loin d’être suffisant pour arriver à des conclusions assurées, et quoique en outre le grand écrivain ait mythifié la sous-race nordique de la race caucasoïde, qu’il appelait « race ariane », aux dépens des autres sous-races qui constituent avec celle-ci le fonds génétique de la nation française.
Jules Ferry – ancien ministre de l’instruction publique, ancien président du conseil, fondateur de l’école laïque –, qui avait sans doute lu Gobineau, n’a pas dit autre chose quand il a souligné la grandeur de la colonisation française dans son fameux discours de 1885 à la chambre des députés : « Messieurs, il faut parler plus haut et plus vrai ! Il faut dire ouvertement qu’en effet les races supérieures ont un droit vis-à-vis des races inférieures… Je répète qu’il y a pour les races supérieures un droit, parce qu’il y a un devoir pour elles. Elles ont le devoir de civiliser les races inférieures. »
Il va de soi, encore faut-il le souligner pour prévenir les mauvais procès que de méchantes gens ne manqueraient pas de nous faire, que la reconnaissance de cette inégalité entre les races, qui est un fait, n’implique aucun jugement de valeur et ne justifie nullement des discriminations illégitimes entre les individus ou les catégories d’individus. Pour les nationaux-libéraux, qui sont attachés au principe républicain de l’égalité civique, les discriminations opérées par la puissance publique ne sont légitimes que si elles sont fondées sur la nationalité, donc sur la préférence nationale, d’une part, ou sur les capacités et les mérites individuels, d’autre part.
On doit préciser en outre qu’il ne s’agit ici que de différences moyennes. Il en résulte que de nombreux congoïdes ont un QI supérieur à celui de nombreux caucasoïdes.
La question qui reste ouverte, s’agissant de certaines différences moyennes entre les races, notamment pour le QI, est de savoir si celles-ci sont intrinsèques et liées à la race, autrement dit aux gènes et autres séquences d’ADN propres à la race, ou si ce sont des différences extrinsèques, statistiques, qui résulteraient de gènes non raciaux, associés par les circonstances aux gènes raciaux. Dans certains cas, on peut affirmer qu’un trait physique ou moral est particulier à une race. De même que les congoïdes ont un nez épaté et des cheveux crépus, le cerveau reptilien dont ils sont dotés, selon l’analyse des trois cerveaux de Paul MacLean, est nettement plus performant que celui des autres races.
Les diverses races ont donc des caractéristiques propres qu’elles ne partagent pas avec les autres. Les races ne se réduisent pas à des différences statistiques entre populations. Ce point de vue réductionniste est erroné. Il y a une essence de la race. Il est cependant remarquable que, s’il y a un type alpin, un type méditerranéen, un type nordique, on ne peut pas parler d’un « type caucasoïde ». Ainsi, c’est la sous-race qui constitue l’unité élémentaire de la raciologie.
Il est certain que les gènes raciaux, ceux qui caractérisent la race, ont autant d’effets sur le moral que sur le physique. C’est pourquoi l’identité d’une société, qui est un système bioculturel, est liée à la composition raciale de sa population.
De surcroît, on peut inférer de la sociobiologie d’Edward Wilson, complétée par la théorie du gène égoïste de Richard Dawkins, que les individus de même race éprouvent normalement les uns pour les autres un sentiment de fraternité raciale. La conscience de race est une source d’altruisme, mais elle peut aussi dégénérer en haine raciale. La race est, en ce sens, une coalition de gènes qui favorise la coopération entre les individus de même race, tout en étant susceptible de conduire à un affrontement entre populations de races différentes, lorsque celles-ci cohabitent sur un même territoire, comme on le voit depuis longtemps aux États-Unis d’Amérique, où le lancinant « problème noir » attend toujours une solution. Ainsi, les sociétés multiraciales sont instables. Elles sont des foyers de guerre civile.
Références
- Bednarik (Robert G.), The Domestication of Humans [La domestication des êtres humains], Routledge, 2020.
- Coon (Carleton S.), Racial Adaptations – a study of the origins, nature & significance of racial variations in humans [Adaptations raciales – étude des origines, de la nature et de la portée des variations raciales chez les êtres humains], Nelson Hall, 1982.
- Coon (Carleton S.), « Populations (Human) » [Populations humaines], The new Encyclopædia Britannica, Macropædia, tome 14, Encyclopædia Britannica, Inc., 1977.
- Coon (Carleton S.), avec Hunt, Jr (Edward E.), The Living Races of Man [Les races vivantes de l’homme], Alfred A. Knopf, 1965.
- Coon (Carleton S.), Histoire de l’homme, Calmann-Lévy, 1958.
- Coon (Carleton S.), Garn (Stanley M.), Birdsell (Joseph B.), Races… a study of the problems of race formation in man [Races… Étude des problèmes de la formation des races chez l’homme], Greenwoof Press, 1950.
- Darlington (Cyril D.), The evolution of man and society [Évolution de l’homme et de la société], Allen and Unwin, 1969.
- Darlington (Cyril D.), Genetics and Man [La génétique et l’homme], Penguin, 1966.
- Darwin (Charles), On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life [De la genèse des espèces sous l’effet de la sélection naturelle, ou conservation des races avantagées dans la lutte pour la vie], John Murray, 1859.
- Dawkins (Richard), Le gène égoïste, Odile Jacob, 2021.
- Gobineau (Arthur de), Essai sur l’inégalité des races humaines, in Œuvres, tome I, Gallimard, coll. « La Pléiade », 1983.
- Hébert (Jean-Pierre), Race et intelligence, Copernic, 1977.
- Herrnstein (Richard), Murray (Charles), The bell curve. Intelligence and class structure in American life [La courbe en cloche. Intelligence et structure de classes dans la société américaine], The Free Press, 1994.
- Jensen (Arthur R.), The g factor [Le facteur g], Greenwood Publishing Group,1998.
- Jensen (Arthur R.), Educability and group differences [Éducabilité et différences entre groupes], Methuen, 1973.
- Jensen (Arthur R.), Genetics & education [Génétique et éducation], Methuen, 1972.
- Lesquen (Henry de) et le Club de l’Horloge, La politique du vivant, Albin Michel,1979.
- Levin (Michael), Why race matters [Pourquoi la race importe], New Century Books, 2016.
- Lynn (Richard), Race differences in intelligence: an evolutionary analysis [Les différences d’intelligence entre les races : analyse évolutionniste], Washington Summit Publishers, 2006.
- Rushton (J. Philippe), Race, evolution, and behavior: a life history perspective [Race, évolution et comportement : vue perspective de l’histoire de la vie], The Charles Darwin Research Institute, 2000.
- Sarich (Vincent), Miele (Frank), Race: the reality of human differences [Race : réalité des différences humaines], Basic Books, 2004.
- Vallois (Henri), Les races humaines, PUF, coll. « Que sais-je ? », 1976.
- Wallace (Alfred), On the tendency of varieties to depart indefinitely from the original type, 1858 ; traduction, « De la tendance des variétés à s’écarter indéfiniment du type primitif », in La sélection naturelle, Elibron Classics, 2006.
- Wilson (Edward), La sociobiologie, éditions du Rocher, 1987.
- Wolpoff (Milford) et Caspari (Rachel), Race and human evolution [Race et évolution de l’homme], Westview Press, 1998.



